
Tricherie
La tricherie est partout, y compris dans la Saintonge moyennageuse : on pèse les âmes, mais Satan essaie de tricher :


Alain Lenoir mis à jour 19-Avr-2019
L'évolution a favorisé de nombreux typre d'interactions entre individus de la même espèce ou d'espèces différentes. Le degré d'interaction varie du simple parasitisme à la coopération (mutualisme). La persistance de la coopération est un paradoxe car la sélection naturelle devrait favoriser les individus qui exploitet leur partenaire (on parle alors de tricherie - cheating) et obtiennent des bénéfices sans fournir en échange une récompense, et cela entraîne une rupture de la coopération. Pour prévenir cela, des mécanismes vont limiter les effets des tricheurs et ainsi permettre le maintien de la coopération. Ces conflits vont se retrouver à tous les niveaux d'organisation, depuis les gènes à l'intérieur d'une cellule, jusqu'aux sociétés (Foster et Kokko 2006). La plupart des travaux portent sur le maintien de la coopération à l'intérieur d'une même espèce par des mécanismes comme la sélection de parentèle, mais le fait d'aider des individus d'une autre espèce est aussi étudié dans de nombreux cas.
La
tricherie intraspécifique
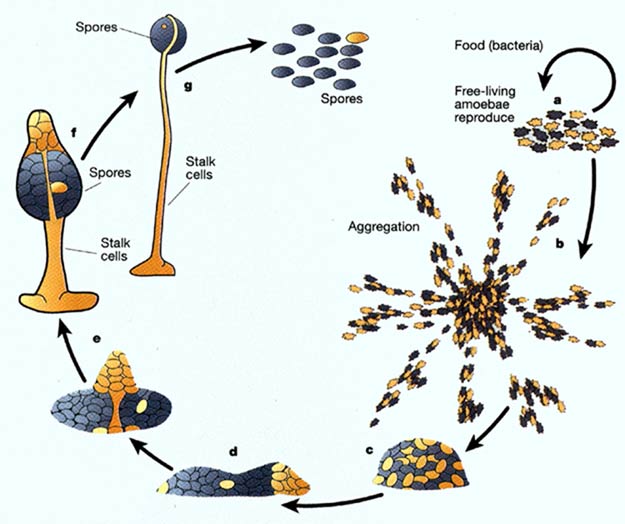
Un exemple bien connu est celui de l'amibe sociale Dictyostelium
discoideum. Elles sont clonales (coefficient de parentèle r
= 1) et forment de très grandes unités comme par exemple celle
que l'on a découvert dans une prairie du Texas. Quand les amibes ne trouvent
plus leur alimentation en bactéries elles s'agrègent en une sorte
de limace multicellulaire. Les limaces se transforment en fructification de
spores. Les cellules de la base (25% des cellules) vont mourir par apoptose
(mort cellulaire programmée) ce qui permet aux autres de devenir des
spores résistantes. Au laboratoire, quand on mixte deux lignées,
l'une d'entre elles se transforme en tricheurs égoïstes qui préfèrent
prendre les meilleures places dans le pédoncule. Les tricheurs ne peuvent
envahir la population car ils ont besoin des non-tricheurs altruistes. En réalité
le conflit est souvent limité dans la nature car les souches sont capables
de discriminer les apaprentés des non-apparentés et vont s'associer
avec les premiers pour éviter la tricherie (Mehdiabadi and al. 2006).
Un comportement de fructification semblable existe aussi chez les bactéries
Myxococcus avec de la tricherie (Velicer et al. 2000).
Chez les fourmis. Pristomyrmex punctatus est une espèce parthénogénétique, des reproductrices tricheuses ont aussi été découvertes. Les plus grosses fourmis pondent plus d'oeufs et font la plus grosse partie de la reproduction pour assurer leur propre descendance. Elles forment une lignée génétique isolée différente des autres fourmis. Ces tricheuses ont été découvertes sur le terrain en 1984 et le système semble stable (Dobata et al. 2009). Les auteurs n'hésitent pas à parler de "cancer social" (Dobata et al 2011).

Pour notre part, nous
avons découvert une forme de tricherie chez des fourmis non-parthénogénétiques,
Aphaenogaster senilis.
Les ouvrières sont capables de pondre des oeufs haploïdes en absence
de la reine, mais certaines échappent à l'inhibition royale et
pondent même en présence de celle-ci. Il existe aussi un mécanisme
de régulation puisque ces ouvrières sont reconnues par les non-tricheuses
car elles ont une odeur cuticulaire un peu différente et elles sont attaquées,
leurs oeufs sont mangés. On parle de "police" dans la colonie
(Ichinose and Lenoir 2009).
L'existence d'un mécanisme de reconnaissance des tricheurs est discuté
par Chapuisat (2009). Ce phénomène a été trouvé
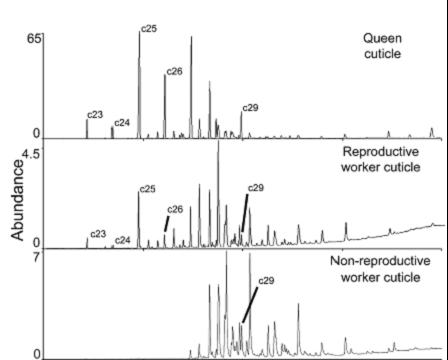
aussi chez Aphaenogaster cockerelli où les tricheuses n'ont
pas le même profil d'hydrocarbures cuticulaires (Smith et al. 2008, 2009)
Voir le profil des A. cockerelli.
Agression des ouvrières
A. senilis pondeuses 
Il est probable que d'autres cas de tricherie seront découverts chez les fourmis, cela dépend un peu si ce sujet est à la mode. C'est ainsi que Cerapachys biroi pratique le policing envers les pondeuses qui trichent (Teseo et al. 2013).
La tricherie intraspécifique existe aussi chez les abeilles avec des ouvrières "anarchistes" (Oldroyd e al 1999, Martin et al 2004).
La tricherie interspécifique
Poissons nettoyeurs. Elle est bien connue chez les vertébrés, par exemple chez les poissons nettoyeurs des récifs coraliens. Ces poissons nettoient d'autres espèces pour éliminer des parasites sur la peau de leur clients. Mais certains nettoyeurs vont aller directement se nourrir du mucus de l'hôte au lieu de s'ocuper des parasites, profitant de la tranquilité près de l'hôte. Comme ailleurs, un mécanisme va limiter la prolifération des tricheurs car les clients observent les nettoyeurs et vont apprendre à les reconnaître et ensuite à les éviter. C'est un processus de score visuel (image-scoring) (Bshary et Grutter 2006). Mais les cheaters vont aussi utiliser l'altruisme avec des clients plus petits en stimulant la zone dorsale de leur client pour améliorer leur image et tromper le client (Bshary 2002).
Les poissons nettoyeurs Garra rufa (Cyprinidae, appelé “the doctor fish”) en Turquie sont utilisés en thérapie humaine pour le psoriasis. Dan son habitat naturel G. rufa broute sur les plantes qui portent des chrysophytes et phytoplancton (Yalcin-Ozdilek and Ekmekci 2006). Avec des pieds humains ils rapent et polissent la peau, ce qui est intéressant pour la nettoyer (Larrochelle 1010).



Nous avons
vu un magasin avec ces poissons à Belgrade en décembre 2016

Poissons
coucous
Il existe des tricheurs chez les poissons du lac Tanganyika qui couvent leurs
oeuf dans leur bouche. Un parasite vient pondre ses oeufs sur ceux de l'hôte
qui les récupère, ce qui réduit considérablement
la survie du squatté. Pourtant celui-ci apprend à déceler
les oeufs étrangers et les écarter.. (voir Herzberg 2018).
Chez les fourmis. Un comportement de nettoyage a été découvert chez des fourmis Dorymyrmex qui lèchent une grande fourmi moissoneuse Pogonomyrmex maricopa dans le désert d'Arizona. On suppose que le bénéfice pour le client est prophylactique, même si cela n'a pas été démontré (Moffett 2010).

Nous avons étudié un coléoptère myrmécophile Sternocoelis hispanus, qui vit dans les nids de fourmis Aphaenogaster en utilisant du mimétisme chimique. Ils ont la même odeur que leur hôte qui les tolère donc. Nous avons observé qu'ils passent un temps considérable sur les ouvrières hôte à les lécher. C'est un autre exemple de nettoyage interspécifique (Lenoir et al 2012). Voir plus largement Lenoir 2013 sur la tricherie chimique.
Voir
- Herzberg, N. (2018). Un coucou dans le lac Tanganyika. Le Monde Science et
Médecine 9 mai 2018. p. 8.
- Larrochelle,
J.-J. (2010). Les bienfaits du "poisson docteur". Le Monde 26 mars.
p. 22.
Publications
- Bshary, R. (2002). Biting cleaner fish use altruism to deceive image-scoring
client reef fish. Proceeding of the Royal Society of London B 269: 2087-2093.
- Bshary, R. and A. S. Grutter (2006). Image scoring and cooperation in a cleaner
fish mutualism. Nature 441: 975-078.
- Chapuisat, M. (2009). Social Evolution: The Smell of Cheating. Current Biology
19(5): R196-R198.
- Dobata, S., T. Sasaki, H. Mori, E. Hasegawa, M. Shimada and K. Tsuji (2009).
Cheater genotypes in the parthenogenetic ant Pristomyrmex punctatus. Proceeding
of the Royal Society of London B 276: 567-574.
- Dobata, S., M. Sasaki, H. Mori, E. Hasegawa, M. Shimada and K. Tsuji (2011).
Persistence of the single lineage of transmissible "social cancer"
in an asexual ant. Molecular Ecology 20(3): 441-455.
- Foster, K. R. and H. Kokko (2006). Cheating can stabilize cooperation in mutualisms.
Proceedings of the Royal Society B-Biological Sciences 273(1598): 2233-2239
- Ichinose, K. and A. Lenoir (2009). Reproductive conflict between laying workers
in the ant Aphaenogaster senilis. Journal of Ethology 27: 475-481. Pdf
- Lenoir, A., Q. Chalon, A. Carvajal, C. Ruel, Á. Barroso, T. Lackner
and R. Boulay (2012). Chemical integration of myrmecophilous guests in Aphaenogaster
ant nests. Psyche Article ID 840860: 12 pages. doi:10.1155/2012/840860. Pdf
- Lenoir, A. (2013). Communication et fraude chimique chez les fourmis. Découverte
388: 24-35. Pdf
- Martin, S. J., N. Châline, B. P. Oldroyd, G. J. Jones and L. W. F. Ratnieks
(2004). Egg marking pheromones of anarchistic worker honeybees (Aps mellifera).
Behavioral Ecology 15: 839-844.
- Mehdiabadi, N. J. and e. al. (2006). Kin preference in a social microorganism.
Nature 442: 881-882.
- Moffett, M. (2010). Adventures Among Ants, California University Press.
- Oldroyd, B. P., L. Halling and T. E. Rinderer (1999). Development and behaviour
of anarchistic honeybees. Proceeding of the Royal Society of London 266: 1875-1878.
- Smith, A., B. Hölldobler and J. Liebig (2008). Hydrocarbon Signals Explain
the Pattern of Worker and Egg Policing in the Ant Aphaenogaster cockerelli.
Journal of Chemical Ecology 34(10): 1275-1282.
- Smith, A. A., B. Hölldober and J. Liebig (2009). Cuticular Hydrocarbons
Reliably Identify Cheaters and Allow Enforcement of Altruism in a Social Insect.
Current Biology 19(1): 78-81.
- Teseo, S., Daniel J. C. Kronauer, P. Jaisson and N. Châline (2013).
Enforcement of Reproductive Synchrony via Policing in a Clonal Ant. Current
Biology(0). http://dx.doi.org/10.1016/j.cub.2013.01.011
- Velicer, G. J., L. Kroos and R. E. Lenski (2000). Developmental cheating in
the social bacterium Myxococcus xanthus. Nature 404: 598-601.
- Yalcin-Ozdilek, S. and F. G. Ekmekci (2006). Preliminary data on the diet
of Garra rufa (Cyprinidae) in the Asi basin (Orontes), Turkey. Cybium 30(2):
177-186.
{kind=link}